

兰科以形态多样丰富多彩的花闻名,尤其是一片高度特化的花被器官——唇瓣,在颜色、形状和结构上极为多样,是吸引传粉者、促进物种多样化的关键(图1)。但兰科唇瓣是如何演化的以及哪些基因可能参与了调控?这些问题一直困扰着进化生物学家。
近日,我院张国进副教授团队与美国宾夕法尼亚州立大学马红教授团队合作在《Plant Communications》上发表了一项大规模研究,通过709种兰花的系统发育基因组学和分析8种兰花的花器官比较转录组,为认识兰花复杂花型的演化机制提供了全新的分子证据。
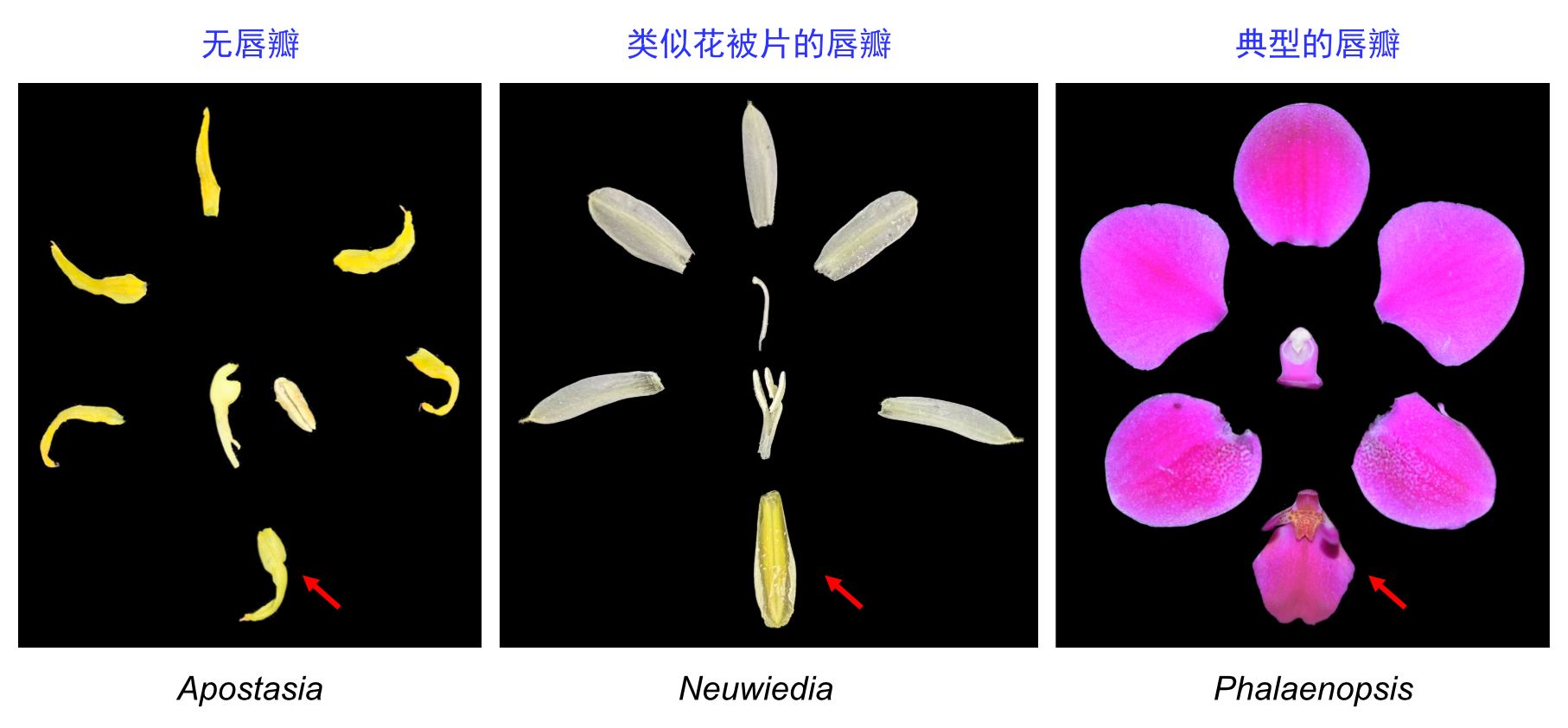
图1 兰科三个代表物种的花结构,分别展示无唇瓣(左)、类似花被片的唇瓣(中-箭头)、典型的唇瓣(右-箭头)的花。
研究首先构建了目前为止采样最广的兰花系统发育基因组学树。通过祖先状态重建发现,最早的兰花花朵其实比较“朴素”,拥有六枚花被片形态相似,没有明显的唇瓣,类似于我们今天看到的百合花。典型唇瓣是在兰科演化的后期,在四个主要亚科的共同祖先中独立起源的。与此同时,侧萼片也开始出现不对称,共同奠定了兰花两侧对称花型的基础(图2)。通过对8种兰花(涵盖四个亚科)的叶片、萼片、花瓣、唇瓣、蕊柱、花药和子房进行转录组测序,研究人员发现:在基因表达层面,唇瓣与普通花瓣最为相似,这为“唇瓣是由花瓣演化而来”的假说提供了分子支持。同时,不同兰花的唇瓣各自积累了大量的特异表达基因,这解释了为何不同兰花的唇瓣形态差异如此之大。

图2 更新的兰科系统发育树以及祖先性状重建识别出的花被器官演化历史;纳入花器官比较转录组分析的物种所在的亚科用星号标出。
经典的MADS-box基因(如AP3和AGL6)在花器官身份决定中发挥核心作用。本研究发现,AP3-2基因在所有兰花的唇瓣中均高表达,功能高度保守。AGL6-2不仅在唇瓣中高表达,还在侧萼片中显著高表达,且这一模式在多个亚科中保守,提示它可能同时调控唇瓣发育和侧萼片的不对称性。研究还发现了一些非MADS-box转录因子家族的新候选调控基因。ALOG家族的G1L基因在唇瓣和蕊柱中高表达,表达模式类似B功能MADS-box基因,可能是唇瓣发育的新调控因子。bZIP6/7基因在蝴蝶兰等晚期分化类群的唇瓣中高表达,可能参与唇瓣后期发育。HD-ZIP家族的HB1c分支基因与黑种草中促进花瓣复杂化的LMI1基因为同源基因,在蝴蝶兰和杓兰等复杂唇瓣中高表达,提示这一调控模式在不同植物中可能被独立招募来驱动花被片复杂化。通过qRT-PCR对蝴蝶兰花发育的三个阶段进行检测,发现G1L在早期高表达,bZIP6/7在中期和晚期表达升高,而部分HD-ZIP基因在早期到中期持续表达。这些结果说明多基因、多阶段协同调控的过程可能促进了兰科花被器官的分化。
这项研究不仅为兰花的系统演化提供了更可靠的框架,也通过大规模的跨物种、器官特异性转录组比较,揭示了兰花复杂花型的可能分子基础(图3)。该研究说明:兰科的花瓣和唇瓣虽然有一定程度的相似性,其实在分子层面已经发生了明显分化,不同兰花类群各自“招募”了一批不同调控基因,可能是唇瓣多样的形态变化的分子基础。这些发现为理解被子植物花器官的多样性演化提供了新视角,也为兰花分子育种提供了宝贵的候选基因资源。

图3 本研究提出的兰科花演化模型。(A)兰科亚科间系统发育关系及纳入比较转录组分析物种(蓝色);带圈数字对应图B中的演化事件;分支标注可能的花被演化事件(见B部分)以及参与演化的候选基因或家族。(B)本研究构建的更综合性的兰科花演化模型,指出了可能的花被演化模式和对称性变化,可能参与调控的基因名称标注在对应的器官上。(C)兰花唇瓣复杂化模式及潜在参与的HD-ZIP基因。
我院张国进副教授为本文的第一作者和共同通讯作者,宾夕法尼亚州立大学马红教授为共同通讯作者,我院硕士研究生黄娜,宾夕法尼亚州立大学研究人员胡宜和博士研究生Clancy A. McIntyre参与了本研究。
(原文信息:Zhang G, Hu Y, Huang N, McIntyre CA, Ma H. Pan-orchid transcriptomics sheds light on candidate regulators and the evolution of complex floral morphology and symmetry.Plant Communications, 2026. Doi:10.1016/j.xplc.2026.101937)